Intracelulárním druhým poslem, zprostředkujícím vazodilatační účinek NO, je cGMP [53, 54, 55, 56]. Bylo popsáno několik mechanismů, jimiž může cGMP působit vazodilataci [243]; jejich skutečný podíl na vazodilataci in vivo většinou není úplně jasný. Jde zejména o snižování citlivosti kinázy lehkého řetězce myosinu ke kalciu a o snižování [Ca2+]i. Na redukci [Ca2+]i se podílí inhibice vápníkových kanálů plasmalemmy i membrány sarkoplasmatického retikula a aktivace pump odčerpávajících Ca2+ z cytoplasmy do extracelulárního prostoru a do sarkoplasmatického retikula. Tyto účinky cGMP jsou zpostředkovány cGMP-dependentní proteinkinázou [243]. Protože kinázy mohou regulovat iontové kanály, a vzhledem k úloze, kterou hrají K kanály v regulaci membránového potenciálu - a v důsledku toho i tenze - plicních cévních myocytů, jsme se rozhodli testovat hypotézu, že NO/cGMP tyto kanály aktivuje a že se tato aktivace významně podílí na mechanismu plicního vazodilatačního účinku NO.
Vliv NO a cGMP na K kanály jsme studovali metodami terčíkového zámku [244]. Podíl jednotlivých typů K kanálů jsme rozlišili pomocí selektivních inhibitorů a biofyzikální charakterizací. Funkčni význam změn aktivity K kanálů jsme studovali na izolovaných kroužcích velkých a periferních plicních tepen in vitro pomocí inhibitorů K kanálů [245, 246, 247, 63, 223].
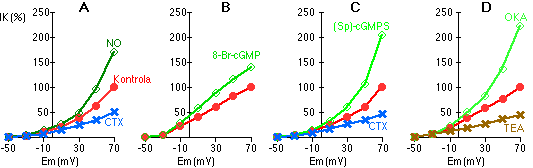
Pokusy s izolovanými plicními cévními myocyty studovanými metodami terčíkového zámku [245, 246] ukázaly, že NO a cGMP (difundující do buňky z měřící pipety anebo podaný externě jako membránově propustný analog) potencovaly draslíkem nesený transmembránový proud (Obrázek 7-1A a B). Tento proud byl inhibovatelný TEA a charybdotoxinem, což ukazuje, že byl zprostředkován KCa kanály (Obrázek 7-1). Buňky, v nichž nad KCa kanály podstatně převládaly 4-AP-senzitivní K kanály, na NO prakticky neodpovídaly [246, 223]. V uspořádání pro měření průchodu proudu jediným kanálem jsme zjistili, že NO rychle a reverzibilně zvyšoval pravděpodobnost otevřeného stavu K kanálů o poměrně velké vodivosti (170 pS), u nichž jsme přímo demonstrovali závislost na kalciu [223]. Skutečnost, že účinek NO na K kanály je zprostředkován cGMP, je potvrzována naším zjištěním, že v přítomnosti inhibitoru guanylátcyklázy, metylénové modři, NO draslíkový proud nezvyšuje [246].
Aktivující vliv NO a cGMP na KCa kanály byl simulován selektivním aktivátorem cGMP-dependentní proteinkinázy (Sp)-guanosin cyklickým 3',5'-fosforothioátem [246, 247] (Obrázek 7-1C). Naopak inhibitor cGMP-dependentní proteinkinázy, látka zvaná H-8, zabránila aktivaci K kanálů oxidem dusnatým [246]. To dohromady ukazuje, že vliv cGMP na KCa kanály je zprostředkován proteinkinázou, tedy fosforylací kanálů. Tomu nasvědčuje i náš nález, že kyselina okadaiková, inhibitor fosfatázy (která defosforyluje proteiny), potencuje proud KCa kanály [246] (Obrázek 7-1D).
Protože poměrně nízká propustnost buněčné membrány je podstatná pro udržování membránového potenciálu, předpokladáli jsme, že aktivace KCa kanálů oxidem dusnatým povede k hyperpolarizaci. Tu jsme také skutečně po aplikaci NO nalezli [223].
Hypotéza, že aktivace KCa kanálů prostřednictvím cGMP (kap. 7.1) je funkčně významnou součástí mechanismu plicní vazodilatační odpovědi na NO, byla testována na izolovaných kroužcích plicních tepen in vitro pomocí inhibitorů K kanálu. Výsledky ukázaly, že relaxace, navozená exogenním NO, je téměř úplně eliminována zvýšením extracelulární koncentrace draslíku [245, 246]. Tento zákrok ruší rozdíl koncentrace K mezi vnitřkem buňky a jejím okolím, který je hnací silou efluxu K při otevření K kanálů. Tento nález je tedy konzistentní s hypotézou, podle níž NO působí vazorelaxaci prostřednictvím otevření K kanálu a následným efluxem K iontů. Identický výsledek jsme získali v pokusech, v nichž byly místo autentického NO použity zaprinast nebo 3-isobutyl-1-metylxantin (IBMX). Zaprinast i IBMX jsou relativně selektivní inhibitory cGMP fosfodiesterázy, která metabolizuje a inaktivuje cGMP. Podobně jako zvýšená extracelulární koncentrace K zablokovaly cGMP-dependentní vazorelaxaci i TEA (relativně selektivní inhibitor KCa kanálů) a charybdotoxin (vysoce selektivní inhibitor KCa kanálů) (Obrázek 7-2). Inhibice K kanálů oslabila cGMP-dependentní vazorelaxaci selektivně, protože vazorelaxační odpověď na papaverin, která není zprostředkována cGMP, nebyla narušena (Obrázek 7-2). Vazorelaxační odpověď na NO byla kompletně zprostředkována cGMP, protože byla zcela zrušena inhibitorem guanylátcyklázy (LY83583) [223].
Tyto výsledky prokazují, že jedním z důležitých mechanismů, jimiž NO působí plicní vazodilataci, je aktivace cGMP-dependentní proteinkinázy, která fosforyluje a tím aktivuje KCa kanály. Výsledný eflux draslíku hyperpolarizuje membránu. Hyperpolarizace inaktivuje napětím řízené vápníkové kanály. V důsledku toho klesá [Ca2+]i a to vede k relaxaci cévního hladkého svalu (Tabulka 7-4). Tento mechanismus však rozhodně nevylučuje současné uplatnění dalších mechanismů, včetně přímého účinku NO (nezprostředkovaného cGMP) na K kanály [248]. Naše výsledky a závěry byly nezávisle potvrzeny jinými autory [249, 250].
Tabulka 7-4: Souhrn výsledků o úloze aktivace KCa kanálu v mechanismu plicní vazodilatační odpovědi na NO
| Hypotéza | Důkaz |
| NO aktivuje guanylátcyklázu | Inhibitory guanylátcyklázy zabrání účinkům NO [246, 223] |
| cGMP prostřednictvím proteinkinázy aktivuje KCa kanály |
NO a cGMP aktivují KCa kanály [245, 246, 223] Aktivátor cGMP-dependentní proteinkinázy [(Sp)-cGMPS] simuluje vliv NO/cGMP na KCa kanály [246, 247] Inhibitor cGMP-dependentní proteinkinázy (H-8) brání aktivačnímu vlivu NO/cGMP na KCa kanály [246] Inhibitor fosfatázy (kyselina okadaiková) simuluje vliv NO/cGMP na KCa kanály [246] |
| Aktivace KCa kanálů působením NO/cGMP hyperpolarizuje cévní myocyty | NO hyperpolarizuje plicní cévní myocyty [223] |
| Aktivace KCa kanálů je podstatná pro vazodilatační odpověď na NO | Inhibitory KCa kanálů zabrání vazodilatační odpovědi na NO [245, 246] |
| Předchozí kapitola |
|
53. Craven PA, DeRubertis FR: Restoration of the responsiveness of purified guanylate cyclase to nitrosoguanidine, nitric oxide, and related activators by heme and heme proteins: evidence for the involvement of the paramagnetic nitrosyl-heme complex in enzyme activation. J Biol Chem 1978; 253: 8433-8443.
54. Gruetter CA, Barry BK, McNamara DB, Gruetter DY, Kadowitz PJ, Ignarro LJ: Relaxation of bovine coronary artery and activation of coronary arterial guanylate cyclase by nitric oxide, nitroprusside and a carcinogenic nitrosoamine. Adv Cyclic Nucleot Res 1979; 5: 211-224.
55. Murad F, Arnold W, Mittal CK, Braughler JM: Properties and regulation of guanylate cyclase and some proposed functions for cyclic GMP. Adv Cyclic Nucleot Res 1979; 11: 175-204.
56. Gruetter CA, Gruetter DY, Lyon JE, Kadowitz PJ, Ignarro LJ: Relationship between cyclic guanosine 3':5'-monophosphate formation and relaxation of coronary arterial smooth muscle by glyceryl trinitrate, nitroprusside, nitrite and nitric oxide: effects of methylene blue and methemoglobin. J Pharmacol Exp Ther 1981; 219: 181-186.
63. Archer SL, Hampl V, Huang JMC, Nelson DP, Tolarova S, Shultz PJ, Weir EK: Nitric oxide and cyclic guanosine monophosphate: effects on potassium channels and vascular tone. In: Weir EK, Archer SL, Reeves JT, Eds. Nitric Oxide and Radicals in the Pulmonary Vasculature. Armonk, Futura, 1996; 391-413.
223. Archer SL, Huang JMC, Reeve HL, Hampl V, Tolarová S, Michelakis E, Weir EK: Differential distribution of electrophysiologically distinct myocytes in conduit and resistance arteries determines their response to nitric oxide and hypoxia. Circ Res 1996; 78: 431-442. (abstrakt)
243. Schmidt HHW, Lohmann SM, Walter U: The nitric oxide and cGMP signal transduction system: regulation and mechanism of action. Biochim Biophys Acta 1993; 1178: 153-175.
244. Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ: Improved patch-clamp techniques for high resolution current recordings from cells and cell-free membrane patches. Pflugers Arch 1981; 391: 85-100.
245. Archer SL, Huang J, Weir EK: Cyclic guanosine monophosphate (c-GMP) and nitric oxide (NO) increase an outward K+ current in rat pulmonary vascular smooth muscle cells. Circulation 1993; 88 (suppl.): I-477.
246. Archer SL, Huang JM-C, Hampl V, Nelson DP, Shultz PJ, Weir EK: Nitric oxide and cGMP cause vasorelaxation by activation of a charybdotoxin-sensitive K channel by cGMP-dependent protein kinase. Proc Natl Acad Sci USA 1994; 91: 7583-7587. (abstrakt)
247. Hampl V, Huang JM, Weir EK, Archer SL: Activation of the cGMP-dependent protein kinase mimics the stimulatory effect of nitric oxide and cGMP on calcium-gated potassium channels. Physiol Res 1995; 44: 39-44. (abstrakt)
248. Bolotina VM, Najibi S, Palacino JJ, Pagano PJ, Cohen RA: Nitric oxide directly activates calcium-dependent potassium channels in vascular smooth muscle. Nature 1994; 368: 850-853.
249. Bialecki RA, Stinson-Fisher C: KCa channel antagonists reduce NO donor-mediated relaxation of vascular and tracheal smooth muscle. Am J Physiol 1995; 12: L152-L159.
250. Paterno R, Faraci FM, Heistad DD: Role of Ca2+-dependent K+ channels in cerebral vasodilatation induced by increases in cyclic GMP and cyclic AMP in the rat. Stroke 1996; 27: 1603-1607.
| Předchozí kapitola |
|