V přítomnosti kyslíku má NO poločas pouhých několik sekund, takže přímé měření jeho množství je technicky poměrně obtížné. Proto většina poznatků o fyziologii NO pochází ze studií využívajících farmakologické inhibice syntézy NO. Nevýhodou tohoto přístupu je nedostatečná specificita inhibitorů NO syntázy (kap. 3.2). Proto jsme považovali za nezbytné pro studium úlohy NO v plicním oběhu pokud možno přímo měřit syntézu NO.
Používali jsme "zlatý standard" analýzy NO, chemiluminescenční metodu [66]. Naše studium této metody a praktické zkušenosti s ní jsou základem našich příspěvků do dvou monografií o metodách měření NO v biomedicínských aplikacích [67, 68]. Podstatou chemiluminescenční metody je vysoce selektivní a rychlá interakce NO s ozónem za vzniku NO2. Některé ze vzniklých molekul NO2 jsou v nestabilním excitovaném stavu a spontánně přecházejí do základního stavu uvolněním fotonu. Tato luminescence je v širokém rozmezí přímo úměrná množství ozónu (které je udržováno stálé) a NO [66]. Výhodou této metody je zejména velmi nízký detekční limit (~1-2 pmol) a možnost měřit NO jak v plynné, tak i v kapalné fázi. Citlivost a přesnost měření v kapalné fázi značně zvyšuje probublávání vzorku konstantním proudem inertního plynu.
Ve vodném prostředí za přítomnosti kyslíku NO rychle oxiduje na NO2, který dále reaguje s NO za vzniku N2O3. Voda reaguje s N2O3 za vzniku nitritů (NO2- ), které v přítomnosti oxyhemoproteinů dále oxidují na nitráty (NO3- ) [69]. Pro sledování rychlosti syntézy NO jsme se proto rozhodli měřit nejen rychlost přírůstku množství NO samotného, ale i přírůstek množství jeho oxidačních produktů. Pomocí silných redukčních činidel (vhodné jsou zejména jodidy nebo vanad) lze v kyselém prostředí za pokojové teploty redukovat NO2- na NO a ten pak detekovat chemiluminescencí [70, 71]. Probíhá-li redukce při teplotách blízkých 100oC, je na NO redukován i NO3- (měření NO3- nebylo ve většině našich pokusů nezbytné, neboť jsme používali média neobsahující oxyhemoproteiny) [69].
Klasickou metodou studia fyziologické úlohy nějaké látky je měření účinku farmakologické inhibice syntézy či účinku této látky na sledovanou fyziologickou funkci. Od začátku devadesátých let jsou k dispozici kompetitivní inhibitory syntézy NO, konkrétně deriváty L-argininu [72, 73]. Mezi nejčastěji používané patří NG-monometyl-L-arginin (L-NMMA), Nomega-nitro-L-arginin (L-NA) a Nomega-nitro-L-arginin metylester (L-NAMA). Deriváty L-argininu však obsahují alkoholové, aminové, anebo karboxylátové skupiny, které mohou interagovat s řadou intracelulárních systémů obsahujících železo [74]. Proto jsme považovali za nezbytné před použitím derivátů L-argininu pro studium úlohy NO v plicní cirkulaci ověřit do té doby běžně přijímaný předpoklad vysoké specificity těchto látek pro inhibici NO syntázy.
K tomuto účelu jsme provedli pokus, v němž jsme současně měřili cévní tonus a tvorbu NO v izolovaných cévách in vitro před a po podání derivátu L-argininu [75]. Použili jsme izolované segmenty (ve tvaru kroužků širokých přibližně 2 mm) aorty a pravé větve plícnice laboratorních potkanů. Cévní kroužky byly mírně nataženy v temperované (37oC) okysličené (PO2=130-140 mmHg) lázni (Earlův roztok) a tenze jejich stěny byla kontinuálně měřena. Obsah NO ve vzorcích lázně, odebraných z blízkosti tkáně, byl měřen chemiluminescenční metodou (kap. 3.1).
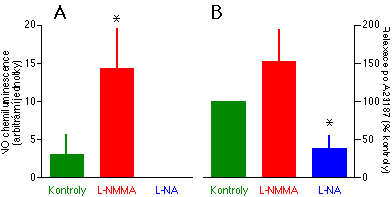
Zjistili jsme, že v rozporu s běžně přijímanou představou byl obsah NO v lázni v blízkosti cévních kroužků signifikantně zvýšen po podání 10-4 M L-NMMA (Obrázek 3-1A). L-NMMA nezabránil vzestupu obsahu NO vyvolanému podáním kalciového ionoforu A23187 5. Vazodilatace vyvolaná v plicních cévách podáním A23187 nebyla oslabená v přítomnosti L-NMMA, spíše měla tendenci ke vzrůstu (Obrázek 3-1B). Obsah NO byl naproti tomu redukován pod detekční limit chemiluminescenčního analyzátoru podáním 2x10-5 M L-NA (Obrázek 3-1A). Vazodilatace vyvolaná podáním A23187 byla signifikantně redukována v přítomnosti L-NA jak v plícnici, tak v aortě (Obrázek 3-1).
Oba studované deriváty L-argininu vyvolaly poměrně slabou konstrikci cévních kroužků (cca 10%). Byla-li však tenze kroužků nejprve i jen velmi mírně zvýšena podáním nízké dávky fenylefrinu (10-8 M), měly deriváty L-argininu poměrně značný vazokonstrikční účinek (>30%). Tomuto vzrůstu tenze nezabránilo odstranění endotelu [75].
Tyto pokusy ukázaly, že L-NMMA není vhodným inhibitorem syntézy NO v plicních cévách , protože tam naopak působí paradoxní zvýšení syntézy NO. Již dříve bylo publikováno, že endoteliální buňky mají schopnost metabolizovat L-NMMA na L-citrulin, který pak může být konvertován na prekurzor oxidu dusnatého L-arginin [80]. Naše studie jako první přímo potvrdila, že L-NMMA může zvyšovat syntézu NO. Skutečnost, že schopnost L-NMMA omezit NO-dependentní vazodilataci je mnohem menší v plicních než v systémových cévách [81, 75], může naznačovat, že plicní endotel metabolizuje L-NMMA na NO snáze než endotel systémových cév. Tato možnost je podporována naším nálezem, že za přítomnosti L-NMMA je množství NO v blízkosti kroužků z plícnice signifikantně vyšší než u kroužků aortálních.
Dalším důležitým nálezem této studie je nespecificky (tedy s inhibicí syntézy NO nesouvisející) vazokonstrikční účinek L-NMMA a L-NA. Zvýšení tonu všech studovaných cév podáním L-NMMA, zejména po jejich prekonstrikci fenylefrinem, nelze vysvětlit odstraněním vazodilatačního působení NO, neboť L-NMMA syntézu NO nejen nesnížil, nýbrž naopak zvýšil. Tento argument neplatí pro L-NA, který syntézu NO lehce snížil. Pro závěr, že část vazokonstrikčního účinku L-NA je nespecifická, však svědčí skutečnost, že tomuto účinku nezabránilo odstranění endotelu, který je výhradním zdrojem NO ve zdravých plicních cévách. Popis nespecifických účinků inhibitorů syntézy NO podtrhuje nutnost nespoléhat při studiu úlohy NO v plicní cirkulaci pouze na použití těchto látek, nýbrž použít i co možná nejpřímější měření syntézy NO. Náš nález nespecifických účinků inhibitorů NO syntázy byl opakovaně potvrzen různými metodami [82, 74, 83, 84].
| Předchozí kapitola |
|
66. Fontijn A, Sabadell AJ, Ronco RJ: Homogenous chemiluminescent measurement of nitric oxide with ozone. Implications for continuous selective monitoring of gaseous air polutants. Anal Chem 1970; 42: 575-579.
67. Archer SL, Shultz PJ, Warren JB, Hampl V, DeMaster EG: Preparation of standards and measurement of nitric oxide, nitroxyl, and related oxidation products. Methods: A Companion to Methods in Enzymology 1995; 7: 21-34.
68. Hampl V, Walters CL, Archer SL: Determination of nitric oxide by the chemiluminescence reaction with ozone. In: Feelisch M, Stamler JS, Eds. Methods in Nitric Oxide Research. Chichester, Wiley, 1996; 310-318.
69. Bush PA, Gonzalez NE, Griscavage JM, Ignarro LJ: Nitric oxide synthase from cerebellum catalyzes the formation of equimolar quantities of nitric oxide and citrulline from L-arginine. Biochem Biophys Res Commun 1992; 185: 960-966.
70. Cox RD: Determination of nitrate and nitrite at the parts per billion level by chemiluminescence. Anal Chem 1980; 52: 332-335.
71. Braman RS, Hendrix SA: Nanogram nitrite and nitrate determination in environmental and biological materials by vanadium (III) reduction with chemiluminescence detection. Anal Chem 1989; 61: 2715-2718.
72. Rees DD, Palmer RMJ, Hodson HF, Moncada S: A specific inhibitor of nitric oxide formation from L-arginine attenuates endothelium-dependent relaxation. Br J Pharmacol 1989; 96: 418-424.
73. Rees DD, Palmer RM, Schulz R, Hodson HF, Moncada S: Characterization of three inhibitors of endothelial nitric oxide synthase in vitro and in vivo. Br J Pharmacol 1990; 101: 746-752.
74. Peterson DA, Peterson DC, Archer S, Weir EK: The non specificity of specific nitric oxide synthase inhibitors. Biochem Biophys Res Commun 1992; 187: 797-801.
75. Archer SL, Hampl V: NG-monomethyl-L-arginine causes nitric oxide synthesis in isolated arterial rings: trouble in paradise. Biochem Biophys Res Commun 1992; 188: 590-596. (abstrakt)
76. Johns A, Lategan TW, Lodge NJ, Ryan US, van Breemen C, Adams DJ: Calcium entry through receptor operated channels in bovine pulmonary artery endothelial cells. Tissue and Cell 1987; 19: 733-745.
77. Peach MJ, Singer HA, Izzo NJ, Loeb AL: Role of calcium in endothelium-dependent relaxation of arterial smooth muscle. Am J Cardiol 1987; 59: A35-A43.
78. Luckhoff A, Pohl U, Mulsch A, Busse R: Differential role of extra- and intracellular calcium in the release of EDRF and prostacyclin from cultured endothelial cells. Br J Pharmacol 1988; 95: 189-196.
79. Blatter LA, Taha Z, Mesaros S, Shacklock PS, Wier WG, Malinski T: Simultaneous measurements of Ca2+ and nitric oxide in bradykinin-stimulated vascular endothelial cells. Circ Res 1995; 76: 922-924.
80. Hecker M, Mitchell JA, Harris HJ, Katsura M, Thiemermann C, Vane JR: Endothelial cells metabolize NG-monomethyl-L-arginine to L-citrulline and subsequently to L-arginine. Biochem Biophys Res Commun 1990; 167: 1037-1043.
81. Archer SL, Cowan NJ: Measurement of endothelial cytosolic calcium concentration and nitric oxide production reveals discrete mechanisms of endothelium-dependent pulmonary vasodilation. Circ Res 1991; 68: 1569-1581.
82. Lippton HL, Hao Q, Hyman A: L-NAME enhances pulmonary vasoconstriction without inhibiting EDRF-dependent vasodilation. J Appl Physiol 1992; 73: 2432-2439.
83. Buxton ILO, Cheek DJ, Eckman D, Westfall DP, Sanders KM, Keef KD: NG-nitro-L-arginine methyl ester and other alkyl esters of arginine are muscarinic receptor antagonists. Circ Res 1993; 72: 387-395.
84. Tseng C-M, Goodman LW, Rubin LJ, Tod ML: NG-monomethyl-L-arginine paradoxically relaxes preconstricted canine intrapulmonary arteries. J Appl Physiol 1993; 74: 549-558.
5 Zvýšení intracelulární koncentrace volného vápníku v endoteliálních buňkách je známým stimulem syntézy NO [76, 77, 78, 79].
| Předchozí kapitola |
|