Plicní oběh je unikátním cévním řečištěm, protože má význam zcela odlišný od ostatních orgánových řečišť. Kyslík v ní není odevzdáván z krve do okolí cév (tkání); naopak, kyslík se v plicní cirkulaci dostává z okolí cév (alveolů) do krve. Tomu odpovídají významné rozdíly funkce a regulace. I nemoci plicního cévního řečiště se odlišuji od patologických stavů, které postihují cévy systémové. Chronické poškození plicních cév se manifestuje plicní hypertenzí. Přestože různé formy plicní hypertenze jsou poměrně časté a jsou spojeny se značnou morbiditou a mortalitou, mechanismus vzniku této poruchy není jasný. Proto jsou i možnosti účinné prevence a léčby plicní hypertenze omezené.
V posledním desetiletí byl objeven biologický význam oxidu dusnatého (NO). Velmi rychle byly nashromážděny důkazy potvrzující, že tato jednoduchá molekula hraje velmi významnou úlohu v řadě fyziologických funkcí, včetně regulace systémového cévního tonu. Bylo přirozené klást si otázku, zda má NO podobně důležitou roli také v regulaci plicních cév, a to jak ve zdraví, tak i při plicní hypertenzi. Tento spis shrnuje naše pokusy, jejichž cílem bylo nalezení odpovědi na tuto otázku.
Plicní cirkulace se v několika ohledech zásadně liší od orgánových cévních rečišť [ref. 1, 2]. To souvisí především s tím, že plicní cévní řečiště samo tvoří celý malý oběh, relativně nezávislý na velkém oběhu, který je naopak tvořen souhrnem všech ostatních orgánových rečišť pozn.1. Jedním z nejnápadnějších rozdílů je skutečnost, že tlak krve je v plicnim oběhu přibližně pět až šestkrát nižší než v systémové cirkulaci [1, 2]. Vysoký tlak ve velkém oběhu je důležitý pro efektivní transport okysličené krve na velké vzdálenosti i proti působení gravitace. V malém oběhu, kde jsou vzdálenosti a vliv gravitace menší, není vysoký krevní tlak nutný. Spíše naopak, vysoký tlak by velmi snadno vedl k poškození tenkého alveolokapilárního rozhraní [3]. Tenké alveolokapilární rozhraní je podstatné pro efektivní difúzi kyslíku do krve.
Další důležitý rozdíl mezi malým a velkým oběhem je v průtoku krve jednotlivými orgány. V malém oběhu protéká prakticky celý srdeční výdej jediným orgánem - plícemi, zatímco ve velkém oběhu protéká každým orgánem jen (větší nebo menší) část srdečního výdeje. Rychlost průtoku krve je tedy mnohem vyšší v plicních než v systémových cévách. Na rychlosti průtoku zavisející střižné napětí na povrchu endotelu (které je velmi důležité pro regulaci endotelu a cévního tonu) je proto zřejmě větší v plicních než v systémových cévách (experimentální potvrzení tohoto tvrzení in vivo by však bylo mimořádně obtížné).
Nižší tlak v plicním řečišti je důsledkem nižšího hemodynamického odporu v plicním řečišti ve srovnání s velkým oběhem. Jednou z příčin nízkého plicního cévního odporu je téměř úplná absence bazálniho tonu hladkého svalu plicních cév [4, 5, 6]. Druhým důvodem je to, že plicní cévy mají tenčí stěnu než cévy systémové [1, 7, 8]. Většina zdravých plicních arteriol dokonce má svalovou vrstvu kolem svého obvodu více nebo méně nekompletní a u mnohých zcela chybí [7, 8]. Systémové arterioly mají naproti tomu silnou vrstvu hladké svaloviny, která vykazuje značný bazální tonus. Tlak, průtok a cévní odpor v plicní cirkulaci a jejich vzájemné vztahy a specifika oproti systémové cirkulaci jsou podrobněji diskutovány v naší nedávno publikované přehledné stati [9]. Důvod o tolik nižší bazálni tenze v plicních cévách ve srovnání se systémovými není znám. Po objevu NO jakožto endoteliálního relaxačního faktoru (EDRF) (kap. 1.2) se objevily spekulace o tom, že příčinou by mohla být podstatně vyšší biosyntéza NO v plicních cévách. Testování této hypotézy bylo jedním z cílů pokusů shrnutých v tomto spisu.
Z hlediska tématu práce je také vhodné připomenout, že nervová regulace je v plicní cirkulaci mnohem méně významná než v cirkulaci systémové [10]. Nervová regulace systémových cév je v zájmu meziorgánové distribuce průtoku a udrženi vysokého krevního tlaku. Tyto potřeby v plicní cirkulaci odpadaji, a ta je proto řízena prakticky výhradně na lokální úrovni. Lokální fyziologická regulace průtoku krve plícemi je podřízena optimálnímu transportu plynů v plicích.
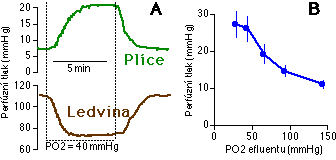
Nejvýznamnějším lokálním mechanismem regulace plicního cévního tonu je hypoxická plicní vazokonstrikce (HPV). Silná vazokonstrikční odpověď na středně silnou hypoxii je charakteristická pro plicní cévy, protože systémová cévní řečiště odpovídají vazodilatací 2 [11, 12] (Obrázek 1-1). Tato dichotomie má důležitý funkční význam. Hypoxická systémová vazodilatace zvyšuje lokální průtok hypoxickým orgánem a tak přizpůsobuje dodávku krve orgánu jeho potřebě kyslíku. HPV naproti tomu omezením perfúze nedostatečně ventilovaných oblastí plic přizpůsobuje průtok krve jednotlivými častmi plic nabídce kyslíku. To je podstatné pro minimalizaci žilní příměsi a tedy i pro optimalizaci okysličení arteriální krve. HPV je navíc klinicky důležitá proto, že při dlouhodobějším působení hypoxie na většinu plicní tkáně může chronická vazokonstrikce přispívat k hypertrofii hladkého svalu cévní stěny a k rozvoji chronické plicní hypertenze (kap. 1.1.2). Mechanismus rozvoje chronické hypoxické plicní hypertenze však není dostatečně znám.
Přestože HPV byla poprvé popsána před více než stoletím [14] a velmi intenzivně studována od konce druhé světové války [15], její mechanismus není dosud jasný. Řada prvků tohoto mechanismu však celkem jasná je (jejich podrobná diskuse je v našem přehledném článku [16]). Je zřejmé, že senzor i efektor HPV jsou lokalizovány především v periferních plicních arteriolách [17, 18, 19, 20, 21]. Od té doby, co tým Dr. Maddenové prokázal hypoxickou kontrakci myocytů izolovaných z periferních plicních arteriol a její absenci u myocytů z plícnice a ze systémových cév [21], je jasné, že mechanismus HPV je inherentní hladkému svalu plicních arteriol. Přitom je však pravděpodobné, že velikost HPV in vivo může být výrazně ovlivňována tkáňovými působky mimosvalového původu [22], zejména z endotelu. Po objevu, že EDRF je NO (kap. 1.2), jehož biosyntéza vyžaduje kyslík, se objevily úvahy, že HPV by mohla být důsledkem hypoxického snížení normálně vysoké biosyntézy tohoto endogenního vazodilatancia. Studium úlohy NO v HPV bylo jednou ze součástí předložené práce (kap. 5).
Plicní hypertenze je syndrom zvýšeného tlaku v plícnici, zvýšeného plicního cévního odporu, nebo obou těchto ukazatelů [23, 24]. Plicní hypertenze je nejčastěji sekundarním onemocněním komplikujícím jiné plicní nebo kardiovaskulární choroby, např. chronickou obstrukční plicní chorobu, mitrální stenózu, či vývojové srdeční vady. Primární plicní hypertenze je poměrně vzácnou, avšak velmi těžkou formou [25, 26]. Plicní hypertenze bez ohledu na příčinu je spojena se značnou morbiditou a mortalitou. Sekundární plicní hypertenze výrazně zhoršuje prognózu primárního onemocnění [24].
Plicní hypertenze klade zvýšené nároky na pravou srdeční komoru, která hypertrofuje přímo úměrně síle plicní hypertenze [27]. Běžné je chronické pravostranné srdeční selhávání, které je také nejčastější příčinou úmrtí [24].
Jedním z nejvýznamnějších faktorů při rozvoji řady forem plicní hypertenze je chronická hypoxie 3. Podle tradiční představy se zde může uplatňovat protahovaná hypoxická plicní vazokonstrikce vedoucí k pracovní hypertrofii cévního hladkého svalu. To vede ke zúžení cévního průsvitu a tedy k růstu cévního odporu. V posledni době je stále zřejmější, že se také podstatným způsobem uplatňuje i přímé působení hypoxie na různé typy buněk, včetně endotelu, vedoucí k proliferaci hladkého svalu a pojivové tkáně cévní stěny. Tyto nedostatečně prozkoumané mechanismy jsou detailněji diskutovány v našich přehledných statích [29, 9].
Po objevu NO jakožto EDRF (kap. 1.2) byly publikovány práce naznačující, že chronická hypoxie snižuje plicní syntézu NO [30, 31]. Byl vysloven názor, že takovéto snížení může být příčinou chronicky zvýšeného plicního cévního tonu a - vzhledem k antiproliferativním účinkům NO [32, 33] - i příčinou proliferace komponent plicní cévní stěny. Studium úlohy endogenního NO při plicní hypertenzi bylo dalším z cílů pokusů popsaných v této práci.
NO je nestabilní radikál, který existuje přednostně v plynné formě. Do nedávné doby byl NO považován pouze za nežádoucí jedovaté znečištění atmosféry, pocházející hlavně ze spalovacích motorů. V průběhu poslední dekády však explozivním tempem přibývalo důkazů o tom, že NO je endogenně produkován v téměř všech savčích tkáních a že hraje celou řadu důležitých biologických úloh, zejména v regulaci hemodynamiky, v zánětu a v neurotransmisi [34, 35, 36, 37, 38, 39, 40, 41]. Přehledný článek o NO v češtině publikoval Prof. Sedláček [42].
Cestu k poznání významu NO v regulaci cévního tonu otevřel zásadní objev Dr. Furchgotta, že pro vazodilatační odpověď na některé agonisty je zcela esenciální přítomnost nepoškozeného endotelu [34], na který se do té doby nahliželo pouze jako na zcela pasivní výstelku cév. Dr. Furchgott také zjistil, že endotel tuto funkci plní prostřednictvím labilního mediátoru, nazývaného endoteliální relaxační faktor (EDRF, endothelium-derived relaxing factor) [34]. V druhé polovině osmdesátých let Dr. Furchgott na základě teoretických úvah a dosavadních poznatků z úsilí o identifikaci EDRF navrhl, že by EDRF mohl být NO. To bylo vzápětí nezávisle experimentálně potvrzeno dvěma laboratořemi [35, 36], i když později bylo upřesněno, že EDRF by mohla být složitější molekula uvolňující NO jako svou aktivní složku [43]. Tato otázka není dosud definitivně dořešena [44], avšak z hlediska hemodynamických účinků NO/EDRF, a tedy této práce, není podstatná.
NO je v savčích tkáních syntetizován z L-argininu a molekulárního kyslíku za vzniku L-citrulinu [45, 46, 47, 48, 49, 50, 51]. Reakce je katalyzována komplexním enzymem, zvaným NO syntáza [49, 52, 51]. Existuje jedna inducibilní forma NO syntázy a dvě formy konstitutivní: endoteliální a mozková. Inducibilní NO syntáza je za normálních podmínek inaktivní a produkuje (relativně neregulovaně) vysoká množství baktericidního NO pouze po aktivaci imunitního systému. Pro regulaci cévního tonu je důležitá endoteliální NO syntáza, která je trvale exprimována. Ta tvorí poměrně menší množství NO, které je však přesně regulováno, zejména intracelulární koncentrací Ca2+ [49, 52, 51].
Intracelulárním druhým poslem, zprostředkujícím vazodilatační účinek NO, je guanosin 3',5' cyklický monofosfát (cGMP) [53, 54, 55, 56]. Mechanismy, jimiž cGMP působí vazodilataci, jsou diskutovány v kapitole 7.
Začátkem devadesátých let, krátce po objevu, že EDRF je NO (nebo látka obsahuící NO jako aktivní skupinu) (kap. 1.2), bylo zjištěno, že intravenózní podání inhibitoru syntézy NO vede k systémové hypertenzi [57, 58, 59, 60]. Tento nález byl interpretován v tom smyslu, že žádoucí cévní tonus a krevní tlak jsou udržovány rovnováhou mezi tonickým vazokonstrikčním působením sympatiku na jedné straně a stálou ("tonickou") endoteliální syntézou vazodilatačního NO na straně druhé [57, 58, 59, 60, 39]. Bylo proto logické předpokládat, že velmi nízký klidový tonus plicních cév může být způsoben vysokou tonickou produkcí NO. Porucha této vysoké bazální produkce NO by pak mohla vést k plicní hypertenzi [30, 31]. Navíc, protože jedním ze substrátů biosyntézy NO je kyslík [48], bylo možno spekulovat o tom, že snížení vysoké bazální produkce NO hypoxií je podkladem HPV. Experimentální testování těchto předpokladů bylo cílem pokusů popsaných v následujícím textu 4. Zajímaly nás konkrétně tyto otázky:
|
|
1. Harris P, Heath D: The Human Pulmonary Circulation (Second edition). Edinburgh, Churchill Livingstone, 1977.
2. Fishman AP: Pulmonary circulation. In: Fishman AP, Fisher AB, Eds. Handbook of Physiology. Section 3: The Respiratory System, Volume I. Circulation and Nonrespiratory Functions. Bethesda, American Physiological Society, 1985; 93-165.
3. West JB, Colice GL, Lee Y-J, Namba Y, Kurdak SS, Fu Z, Ou L-C, Mathieu-Costello O: Pathogenesis of high-altitude pulmonary oedema: direct evidence of stress failure of pulmonary capillaries. Eur Respir J 1995; 8: 523-529.
4. Emery CJ, Bee D, Barer GR: Mechanical properties and reactivity of vessels in isolated perfused lungs of chronically hypoxic rats. Clin Sci 1981; 61: 569-580.
5. Adnot S, Chabrier PE, Brun-Buisson C, Viossat I, Braquet P: Atrial natriuretic factor attenuates the pulmonary pressor response to hypoxia. J Appl Physiol 1988; 65: 1975-1983.
6. Hyman AL, Kadowitz PJ: Tone-dependent responses to acetylcholine in the feline pulmonary vascular bed. J Appl Physiol 1988; 64: 2002-2009.
7. Hislop A, Reid L: Normal structure and dimensions of the pulmonary arteries in the rat. J Anat 1978; 125: 71-83.
8. Reid LM: Structural remodeling of the pulmonary vasculature by environmental change and disease. In: Wagner WW, Weir EK, Eds. The Pulmonary Circulation and Gas Exchange. Armonk, Futura, 1994; 77-110.
9. Herget J, Hampl V: Pulmonary circulation. In: Šrámek BB, Valenta J, Klimeš F, Eds. Biomechanics of the Cardiovascular System. Prague, Czech Technical University Press, 1995; 327 - 336.
10. Barnes PJ, Liu SF: Regulation of pulmonary vascular tone. Pharmacol Rev 1995; 47: 87-131.
11. Hampl V, Weir EK, Archer SL: Endothelium-derived nitric oxide is less important for basal tone regulation in the pulmonary than the renal vessels of adult rat. J Vasc Med Biol 1994; 5: 22-30.
12. Weir EK, Archer SL: The mechanism of acute hypoxic pulmonary vasoconstriction: the tale of two channels. FASEB J 1995; 9: 183-189.
13. Hampl V, Herget J: Acute pneumonia reversibly inhibits hypoxic vasoconstriction in isolated rat lungs. Physiol Res 1992; 41: 147-150.
14. Bradford JR, Dean HP: The pulmonary circulation. J Physiol 1894; 16: 34-96.
15. von Euler US, Liljestrand G: Observations on the pulmonary arterial blood pressure in the cat. Acta Physiol Scand 1946; 12: 301-320.
16. Hampl V, Herget J: Possible mechanisms of oxygen sensing in the pulmonary circulation. Physiol Res 1991; 40: 463-470.
17. Kato M, Staub NC: Response of small pulmonary arteries to unilobar hypoxia and hypercapnia. Circ Res 1966; 19: 426-439.
18. Nagasaka K, Bhattacharya J, Nanjo S, Gropper MA, Staub NC: Micropuncture measurement of lung microvascular pressure profile during hypoxia in cats. Circ Res 1984; 54: 90-95.
19. Madden JA, Dawson CA, Harder DR: Hypoxia-induced activation in small isolated pulmonary arteries from the cat. J Appl Physiol 1985; 59: 113-118.
20. Shirai M, Sada K, Ninomiya I: Effects of regional alveolar hypoxia and hypercapnia on small pulmonary vessels in cats. J Appl Physiol 1986; 61: 440-448.
21. Madden JA, Vadula MS, Kurup VP: Effects of hypoxia and other vasoactive agents on pulmonary and cerebral artery smooth muscle cells. Am J Physiol 1992; 263: L384-L393.
22. Hampl V, Herget J: Tkáňové působky ovlivňující hypoxickou plicní vazokonstrikci. Čs fyziol 1991; 40: 1-23.
23. Reeves JT, Groves BM: Approach to the patient with pulmonary hypertension. In: Weir EK, Reeves JT, Eds. Pulmonary Hypertension. Mount Kisco, Futura, 1984; 1-44.
24. Weir EK, Archer SL, Rubin LJ: Pulmonary hypertension. In: Willerson JT, Cohn JN, Eds. Cardiovascular Medicine. New York, Churchill Livingstone, 1995; 1495-1523.
25. D'Alonzo GE, Barst RJ, Ayres SM, Bergofsky EH, Brundage BH, Detre KM, Fishman AP, Goldring RM, Groves BM: Survival in patients with primary pulmonary hypertension. Results from a national prospective registry. Ann Intern Med 1991; 115: 343-349.
26. Abenhaim L, Moride Y, Brenot F, Rich S, Benichou J, Kurz X, Higenbottam T, Oakley C, Wouters E: Appetite-suppressant drugs and the risk of primary pulmonary hypertension. N Engl J Med 1996; 335: 609-616.
27. Kolář F, Ošťádal B, Procházka J, Pelouch V, Widimský J: Comparison of cardiopulmonary response to intermittent high-altitude hypoxia in young and adult rats. Respiration 1989; 56: 57-62.
28. Reeves JT, Herget J: Experimental models of pulmonary hypertension. In: Weir EK, Reeves JT, Eds. Pulmonary Hypertension. Mount Kisco, Futura, 1984: 361-391.
29. Hampl V: Hypoxická plicní hypertenze. Čs fyziol 1991; 40: 255-271.
30. Adnot S, Raffestin B, Eddahibi S, Braquet P, Chabrier P-E: Loss of endothelium-dependent relaxant activity in the pulmonary circulation of rats exposed to chronic hypoxia. J Clin Invest 1991; 87: 155-162.
31. Dinh-Xuan AT, Higenbottam TW, Clelland CA, Pepke-Zaba J, Cremona G, Butt AY, Large SR, Wells FC, Wallwork J: Impairment of endothelium-dependent pulmonary-artery relaxation in chronic obstructive lung disease. N Engl J Med 1991; 324: 1539-1547.
32. Garg UC, Hassid A: Nitric oxide-generating vasodilators and 8-bromo-cyclic guanosine monophosphate inhibit mitogenesis and proliferation of cultured rat vascular smooth muscle cells. J Clin Invest 1989; 83: 1774-1777.
33. Yang WD, Ando J, Korenaga R, Toyo-oka T, Kamiya A: Exogenous nitric oxide inhibits proliferation of cultured vascular endothelial cells. Biochem Biophys Res Commun 1994; 203: 1160-1167.
34. Furchgott R, Zawadzki J: The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 1980; 288: 373-376.
35. Ignarro LJ, Buga GM, Wood KS, Byrns RE, Chaudhuri G: Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proc Natl Acad Sci USA 1987; 84: 9265-9269.
36. Palmer RMJ, Ferrige AG, Moncada S: Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature 1987; 327: 524-526.
37. Ignarro LJ: Endothelium-derived nitric oxide: actions and properties. FASEB J 1989; 3: 31-36.
38. Moncada S, Palmer RMJ, Higgs EA: Nitric oxide: physiology, pathophysiology, and pharmacology. Pharmacol Rev 1991; 43: 109-142.
39. Moncada S: The 1991 Ulf von Euler lecture: The L-arginine:nitric oxide pathway. Acta Physiol Scand 1992; 145: 201-227.
40. Furchgott RF: The discovery of endothelium-dependent relaxation. Circulation 1993; 87: V-3-V-8.
41. Moncada S, Higgs A: The L-arginine-nitric oxide pathway. N Engl J Med 1993; 329: 2002-2012.
42. Sedláček J: NO: nový typ mezibuněčné komunikace. Čs fyziol 1993; 42: 11-17.
43. Myers P, Minor RJ, Guerra RJ, Bates J, Harrison D: Vasorelaxant properties of the endothelium-derived relaxing factor more closely resemble S-nitrosocysteine than nitric oxide. Nature 1990; 345: 161-163.
44. Feelisch W, de Poel M, Zamora R, Deussen A, Moncada S: Understanding the controversy over the identity of EDRF. Nature 1994; 368: 62-65.
45. Palmer RMJ, Rees DD, Ashton DS, Moncada S: L-arginine is the physiological precursor for the formation of nitric oxide in endothelium-dependent relaxation. Biochem Biophys Res Commun 1988; 153: 1251-1256.
46. Moncada S, Palmer RMJ, Higgs EA: Biosynthesis of nitric oxide from L-arginine. Biochem Pharmacol 1989; 38: 1709-1715.
47. Palmer MJ, Moncada S: A novel citrulline-forming enzyme implicated in the formation of nitric oxide by vascular endothelial cells. Biochem Biophys Res Commun 1989; 158: 348-352.
48. Leone AM, Palmer RM, Knowles RG, Francis PL, Ashton DS, Moncada S: Constitutive and inducible nitric oxide synthases incorporate molecular oxygen into both nitric oxide and citrulline. J Biol Chem 1991; 266: 23790-23795.
49. Nathan C: Nitric oxide as a secretory product of mammalian cells. FASEB J 1992; 6: 3051-3064.
50. Katusic ZS, Cosentino F: Nitric oxide synthase: from molecular biology to cerebrovascular physiology. News Physiol Sci 1994; 9: 64-67.
51. Nathan C, Xie Q-W: Regulation of biosynthesis of nitric oxide. J Biol Chem 1994; 269: 13725-13728.
52. Knowles RG, Moncada S: Nitric oxide synthases in mammals. Biochem J 1994; 298: 249-258.
53. Craven PA, DeRubertis FR: Restoration of the responsiveness of purified guanylate cyclase to nitrosoguanidine, nitric oxide, and related activators by heme and heme proteins: evidence for the involvement of the paramagnetic nitrosyl-heme complex in enzyme activation. J Biol Chem 1978; 253: 8433-8443.
54. Gruetter CA, Barry BK, McNamara DB, Gruetter DY, Kadowitz PJ, Ignarro LJ: Relaxation of bovine coronary artery and activation of coronary arterial guanylate cyclase by nitric oxide, nitroprusside and a carcinogenic nitrosoamine. Adv Cyclic Nucleot Res 1979; 5: 211-224.
55. Murad F, Arnold W, Mittal CK, Braughler JM: Properties and regulation of guanylate cyclase and some proposed functions for cyclic GMP. Adv Cyclic Nucleot Res 1979; 11: 175-204.
56. Gruetter CA, Gruetter DY, Lyon JE, Kadowitz PJ, Ignarro LJ: Relationship between cyclic guanosine 3':5'-monophosphate formation and relaxation of coronary arterial smooth muscle by glyceryl trinitrate, nitroprusside, nitrite and nitric oxide: effects of methylene blue and methemoglobin. J Pharmacol Exp Ther 1981; 219: 181-186.
57. Gardiner SM, Compton AM, Bennett T, Palmer RMJ, Moncada S: Regional haemodynamic changes during oral ingestion of NG-monomethyl-L-arginine or NG-nitro-L-arginine methyl ester in conscious Brattleboro rats. Br J Pharmacol 1990; 101: 10-12.
58. Gardiner SM, Compton AM, Bennett T, Palmer RMJ, Moncada S: Control of regional blood flow by endothelium-derived nitric oxide. Hypertension 1990; 15: 486-492.
59. Gardiner SM, Compton AM, Bennett T, Palmer RMJ, Moncada S: Persistent haemodynamic changes following prolonged infusions of NG-monomethyl-L-arginine in conscious rats. In: Moncada S, Higgs EA, Eds. Nitric Oxide from L-arginine: A Bioregulatory System. Amsterdam, Elsevier, 1990; 489-491.
60. Gardiner SM, Kemp PA, Bennett T, Palmer RMJ, Moncada S: Nitric oxide synthase inhibitors cause sustained, but reversible, hypertension and hindquarters vasoconstriction in Brattleboro rats. Eur J Pharmacol 1992; 213: 449-451.
61. Archer SL, Hampl V, Huang J, Cowan N: The importance of calcium in the regulation of EDRF synthesis in the pulmonary vasculature. In: Weir EK, Hume JR, Reeves JT, Eds. Ion Flux in Pulmonary Vascular Control. New York, Plenum, 1993; 223-246.
62. Archer S, Hampl V, McKenzie Z, Nelson D, Huang J, Shultz P, Weir EK: Role of endothelial-derived nitric oxide in normal and hypertensive pulmonary vasculature. Semin Respir Med 1994; 15: 179-189.
63. Archer SL, Hampl V, Huang JMC, Nelson DP, Tolarová S, Shultz PJ, Weir EK: Nitric oxide and cyclic guanosine monophosphate: effects on potassium channels and vascular tone. In: Weir EK, Archer SL, Reeves JT, Eds. Nitric Oxide and Radicals in the Pulmonary Vasculature. Armonk, Futura, 1996; 391-413.
64. Hampl V, Cornfield DN, Huang J, Shultz PJ, Weir EK, Archer SL: Nitric oxide. In: Peacock AJ, Ed. Pulmonary Circulation: A Handbook for Clinicians. London, Chapman and Hall, 1996; 99-114.
65. Hampl V, Archer SL: The role of endogenous nitric oxide in acute hypoxic pulmonary vasoconstriction. In: Zapol WM, Bloch KD, Eds. Nitric Oxide and the Lung. New York, Marcel Dekker, 1997; 113-135.
|
|
1 V tomto textu, tak jako v příslušné odborné literatuře, se pro řečiště velkého oběhu používá také termín "systémové".
2 Je zde řeč o fysiologicky relevantní hypoxii, tedy takové, která je kompatibilní s přežitím tkáně. Příliš silná hypoxie až anoxie může působit vazokonstrikci některých systémových cév. Navíc v některých případech může být lokálně zprostředkovaná systémová vazodilatace potlačena vazokonstrikčním působením sympatiku.
3 Význam chronické hypoxie při vzniku plicní hypertenze podtrhuje skutečnost, že chronická expozice hypoxii je nejběžnějším experimentálním modelem plicní hypertenze [28].
4 Naše nálezy ohledně úlohy NO v plicní cirkulaci a jejich srovnání s nálezy jiných jsou předmětem několika našich přehledných statí [61, 62, 63, 64, 65].
|
|