Před zahájením mých studií úlohy NO v regulaci plicní cirkulace byly publikovány pouze tři práce zabývající se tímto tématem. Brashersová se spolupracovníky [85] použili k inhibici EDRF kyselinu eicosatetrayenovou a nordihydroguaiareovou a hydrochinon a neshledali signifikantní změnu bazálního tonu v izolovaných plicích potkana. Zásadním problémem této práce, publikované před objevem schopnosti derivátů L-argininu inhibovat NO syntázu, je značná neselektivita použitých inhibitorů (např. inhibice syntézy eikosanoidů a oxidační účinky, což obojí ovlivňuje plicní cévní tonus [86, 87]). Mazmanian a spolupracovníci [88] nenalezli signifikantní zvýšení bazálního tonu izolovaných plic potkana po podání metylénové modři. Jedním z hlavních účinků metylénové modři je inhibice guanylátcyklázy, která zprostředkuje působení NO a řady dalších stimulů na cévní tonus. Archer a spolupracovníci [89] poprvé použili při studiu plicního cévního tonu derivátů L-argininu, konkrétně L-NMMA. Bazální tonus v izolovaných plicích potkana nebyl podáním L-NMMA zvýšen. Tento nález byl v průbehu mojí práce na tomto tématu potvrzen dalšími autory, rovněž s použitím L-NMMA [90, 91]. Společným problémem všech těchto studií je nedostatečná selektivita použitých inhibitorů. Např. nezvýšený bazální tonus po podání L-NMMA [89, 90, 91] má minimální vypovídací schopnost o úloze NO vzhledem k našemu následnému zjištění, že L-NMMA může ve skutečnosti syntézu NO potencovat [75] (kap. 3.2). Proto jsme se rozhodli tuto otázku důkladně prověřit pomocí několika různých inhibitorů NO syntázy a přímým měřením tvorby NO. Testovanou hypotézou bylo, že v plicní cirkulaci - na rozdíl od systémových cév - neexistuje funkčně významná bazální syntéza NO.
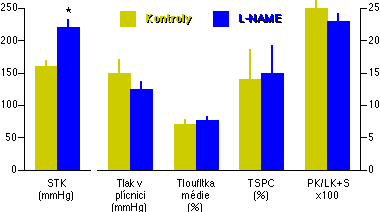
V první z těchto studií [92] jsme se zaměřili na chronickou inhibici NO syntázy. Vycházeli jsme z představy, že je-li klidová syntéza NO v plicních cévách důležitá pro udržování nízkého plicního cévního odporu, pak by její dlouhodobá inhibice měla vyvolat známky plicní hypertenze. Laboratorním potkanům byl do vody na pití přidáván L-NAME (1.85 mM) a po třech týdnech byla měřena systémová a plicní hemodynamika a závislost mezi tlakem a průtokem v izolovaných plicích.
Použitá dávka L-NAME byla dostatečná k vyvolání signifikantní systémové hypertenze (Obrázek 4-1), což potvrdilo přítomnost inhibice NO syntázy. Tlak v plícnici však nebyl vyšší než u kontrol, naopak, byla zde zřetelná tendence k poklesu (Obrázek 4-1), zřejmě v souvislosti se sníženým srdečním výdejem. L-NAME posunul vztah mezi perfúzním tlakem a průtokem v izolovaných plicích k poněkud vyšším tlakům, avšak podstatně méně než klasický stimulus pro plicní hypertenzi, chronická hypoxie. Zvýšená muskularizace periferních plicních cév, typická pro plicní hypertenzi [93, 8], nebyla po chronické inhibici NO syntázy přítomná. Nebyla zvýšená ani tloušťka médie muskularizovaných plicních cév, ani podíl počtu muskularizovaných cév na celkovém počtu periferních plicních cév (Obrázek 4-1). Pravolevý srdeční index, zvýšený při plicní hypertenzi, nevzrostl během dlouhodobé inhibice NO syntázy (Obrázek 4-1).
Regresní analýza individuálních hodnot srdečního výdeje a tlaku v plícnici ukázala, že hodnoty ležely na regresních přímkách závislosti tlaku a průtoku, které byly u kontrol a po chronické inhibici NO syntázy prakticky totožné [65 - obr. 5].
Tyto výsledky tedy ukazují, že dlouhodobá inhibice NO syntázy působí významnou systémovou, avšak nikoliv plicní, hypertenzi. To je v souladu s naší pracovní hypotézou, podle níž v plicní cirkulaci - na rozdíl od cirkulace systémové - neexistuje funkčně významná bazální syntéza NO.
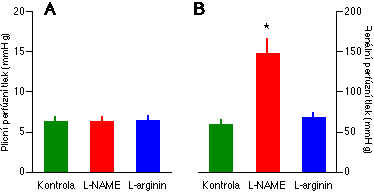
Představu, že bazální syntéza NO je podstatně méně významná v plicních než v systémových cévách jsme podrobněji testovali v další studii [94, 11]. V ní jsme přímo srovnáváli účinek dvou inhibitorů NO syntázy (L-NA a L-NAME) na současně studované plicní a systémové cévní řečiště izolované z téhož laboratorního potkana. Jako modelové systémové cévní řečiště jsme použili řečiště renální. Pro tuto studii jsme vyvinuli nový preparát, ledvinu a ventilované plíce, izolované z téhož potkana a koperfundované ex vivo společným perfuzátem. Jedná se v podstatě o kombinaci a modifikaci klasických preparátů izolovaných plic [95, 96, 97] a izolované ledviny [98, 99]. V zájmu prevence ischémie obou organů je však chirurgická příprava preparátu mnohem obtížnejší než izolace každého orgánu zvlášť. Plíce slouží jako oxygenátor renálního perfuzátu. Perfuzní okruh je připraven tak, že část perfuzátu vytékajícího z plic je čerpána do ledvin (průtok plícemi je vyšší než průtok ledvinou; průtok oběma orgány je konstantní). Renální žíla je přeťata, ne však nakanylována, a výtok z ní volně odchází do malé Petriho misky, v níž je ledvina umístěna, a tak udržuje ledvinu ve vlhkém prostředí. Přebytek tekutiny v Petriho misce volně odtéká do hlavního rezervoáru perfuzátu. Moč je odváděna nakanylovaným močovodem. Toto uspořádání vylučuje variabilitu způsobenou použitím rozdílných zvířat a experimentálních podmínek pro studium regulace plicního a systémových cévních řečišť.
Životnost preparátu potvrdila přítomnost opačných odpovědí na hypoxii (v souladu se situací in vivo): vazodilatace v ledvině a vazokonstrikce v plicích (Obrázek 1-1). Plíce tolerovaly prakticky bez jakéhokoliv zvýšení perfúzního tlaku (který při konstantním průtoku přímo odráží cévní odpor) dávky L-NA i L-NAME, které v ledvinách způsobily dvojnásobné zvýšení tlaku (Obrázek 4-2). Vztah tlaku a průtoku v ledvinách byl po podání L-NA i L-NAME podstatně strmější než za kontrolních podmínek, zatímco v plicích nebyl tento vztah deriváty L-argininu ovlivněn. To naznačuje, že v plicích buď neexistuje, nebo se funkčně neuplatňuje, syntéza NO vyvolaná zvýšenim průtoku, která je běžná v cévách systémových [100, 101, 102].
Vazokonstrikční odpověď plic i ledvin na angiotensin II byla signifikantně zvýšena oběma inhibitory NO syntázy. Například v plicích zvýšil L-NA přírůstek perfúzního tlaku způsobený angiotensinem II ze 3.0 +/- 0.4 na 11.8 +/- 2.5 mmHg. Nejpravděpodobnějším mechanismem je zábrana syntézy NO vyvolané vazokonstrikcí [91]. To ukazuje, že dávka derivátu L-argininu byla pro inhibici dostatečná a že ledviny neuvolňovaly do perfuzátu žádnou (neznámou) látku, která by účinku L-NA a L-NAME v plicích bránila. Účinky kompetitivních inhibitorů NO syntázy bylo možno zvrátit podáním vyšší dávky substrátu NO syntázy L-argininu (Obrázek 4-2). Tato skutečnost, společně s použitím dvou různých inhibitorů, značně snižuje pravděpodobnost, že by naše nálezy byly důsledkem nespecifických účinků inhibitorů NO syntázy.
Už před publikací této naší studie existovaly práce popisující renální vazokonstrikční odpověď na deriváty L-argininu [103, 104, 105, 106, 107, 108, 109, 99]. V průbehu naší práce na této studii přibývalo publikací (včetně našich vlastních: kap. 4.1 a kap. 4.3) popisujících absenci plicní vazokonstrikční odpovědi na inhibici NO syntázy [89, 90, 110, 91, 111, 112, 113, 92, 114, 115, 116, 117, 118, 119, 120]. Tato naše studie je však prioritní v tom, že účinek inhibitorů NO syntázy přímo srovnává mezi těmito dvěma orgány s vyloučením variability mezi laboratořemi, druhy, kmeny, jedinci a experimentálními podmínkami. Ovlivnění vztahu mezi renálním perfúzním tlakem a průtokem inhibicí NO syntázy nebylo před touto studií studováno.
Zajímavým poznatkem této studie je také skutečnost, že inhibice NO syntázy nezmenšila hypoxickou renální vazodilataci. Renální perfúzní tlak se snížil o 48 +/- 6 mmHg před podáním L-NAME a o 54 +/- 5 mmHg po něm (P>0.05). To naznačuje, že NO se neúčastní mechanismu hypoxické dilatace systémových cév. Tento závěr byl později potvrzen pro kosterní sval [121]. Existují však i údaje svědčící pro opak [122, 123].
Měření hemodynamických důsledku farmakologické inhibice NO syntázy sleduje spíše funkční úlohu NO než přímo úroveň syntézy NO jako takovou a navíc je zatíženo možností zkreslení nespecifickými účinky inhibitorů (kap. 3.2). Kromě toho je možné, že za normálních podmínek je bazální tonus plicních cév prakticky nulový i bez přispění NO. Pak by i vysoká syntéza NO nemohla způsobit další vazodilataci, a tedy ani inhibice endogenní syntézy NO by se nemusela projevit vazokonstrikcí.
Z těchto důvodů jsme se rozhodli měřit syntézu NO v izolovaných plicích přímějším způsobem [119]. Pomocí redukce s jodidem draselným v kyselém prostředí jsme konvertovali konečný oxidační produkt NO v umělém perfuzátu, NO2-, zpět na NO 6. Ten jsme pak (spolu s tou částí NO, která před odběrem nezoxidovala) detekovali chemiluminescencí (kap. 3.1). Jedno měření obsahu NO+NO2- v perfuzátu opouštějícím izolované plíce potkana bylo provedeno na začátku perfúze. Bylo zjištěno, že výsledek se neliší od identického měření perfuzátu před zahájením perfúze (tj. měření v perfúzním roztoku, který se ještě nedostal do kontaktu s tkání). Měření pak bylo opakováno po hodině perfúze. Od této hodnoty byla odečtena hodnota naměřená na začátku perfúze a výsledek tedy odpovídal množství NO uvolněného plícemi do perfuzátu za hodinu. U separátní skupiny izolovaných plic bylo chemiluminescencí měřeno množství NO ve vydechovaném vzduchu.
Zjistili jsme [119], že množství NO uvolněného do perfuzátu během jedné hodiny (20 +/- 190 pmol, n = 6) nebylo statisticky významně odlišné od nuly. Přitom schopnost této metody detekovat endogenní NO byla potvrzena hodnotou zjištěnou v plicích chronicky hypoxických potkanů, která byla statisticky významně vyšší než nula (kap. 6.4). V žádném z našich pokusů nebyl NO detekovatelný ve vzduchu vydechovaném z izolovaných plic potkana [124].
Tyto výsledky tedy poprvé přímo demonstrovaly, že bazální produkce NO je v normální plicní cirkulaci minimální. Tento závěr byl dále potvrzen studií s plicními endoteliálními buňkami v kultuře [125, 126]. Metodika měření NO byla prakticky identická s předchozí studií [119] kromě vylepšení několika technických detailů a kratší doby superfúze buněčné kultury (10 min). Akumulace NO+NO2- v superfuzátu byla za kontrolních podmínek nevýznamná (Obrázek 5-2). Schopnost metody detekovat produkci NO byla potvrzena statisticky významně zvýšenou hodnotou při vystavení buněk akutní hypoxii.
NO má poměrně vysoký rozdělovací koeficient (~20) 7 . Proto je pravděpodobné, že značná část NO uvolněného z tenkostěnných plicních cév by se měla dostat do alveolárního vzduchu a následně být detekována ve vydechovaném vzduchu. Lidský dech skutečně obsahuje NO [127, 128]. Rozhodli jsme se tedy zjistit, zda NO detekovaný v lidském dechu pochází z plicních cév.
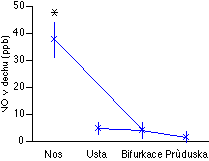
Pomocí chemiluminescence jsme měřili NO ve vzduchu vydechovaném z nosu, úst, průdušnice a distálního bronchu [129]. Zjistili jsme, že koncentrace NO byla největší ve vzduchu vydechovaném nosem, zatímco hodnoty z distálního bronchu byly na mezi detekčního limitu metody (1 molekula NO z každé miliardy molekul) (Obrázek 4-3).
Dýchací cesty jsou poměrně bohaté na NO syntázu [130, 131, 132], takže je pravděpodobné, že většina z minimálního množství NO detekovaného v průduškách pochází spíše z epitelu dýchacích cest než z endotelu plicních cév. Koncentrace NO ve vydechovaném vzduchu byla navíc signifikantně potlačena podáváním kortikosteroidů, což ukazuje, že značná část vydechovaného NO je tvořena inducibilní formou NO syntázy. Pouze inducibilní forma je totiž inhibována steroidy [51]. Inducibilní NO syntáza se uplatňuje při obraně proti infekci, zatímco NO regulující cévní tonus je syntetizován konstitutivní formou NO syntázy. I tento nález tedy nepřímo potvrzuje, že NO v lidském dechu nepochází z endotelu plicních cév. Skutečnost, že naprostá většina NO v lidském dechu pochází z nosu byla opakovaně potvrzena [133, 134, 135, 136, 137] a je dnes více méně všeobecně přijímána. Zajímavý je koncept, podle něhož většina tohoto NO pochází z paranasálních sínů, kde zajišťuje jejich obranu proti infekci [131]. Závěr, že NO v dechu nepochází z plicní cirkulace, potvrzují i naše pokusy s izolovanými plícemi potkanů (kde chybí horní dýchací cesty). Tam je NO konzistentně nedetekovatelný [124].
Výsledky popsané v kapitole 4 (shrnuje je Tabulka 4-1) dohromady ukazují, že bazální syntéza NO je ve zdravých plicních cévách minimální a funkčně nevýznamná. Tím se plicní cirkulace nápadně odlišuje od cirkulace systémové, kde je tonická produkce NO významná pro nastavení bazální cévní tenze.
| 1. | Dlouhodobá inhibice NO syntázy působí významnou systémovou, avšak nikoliv plicní, hypertenzi. |
[92]
|
| 2. | Dávka inhibitorů NO syntázy, která zdvojnásobila cévní odpor v izolované ledvině, neměla žádný vliv na cévní odpor v izolovaných plicích. |
[11]
|
| 3. | NO ani jeho oxidační produkt NO2- není detekovatelný v perfuzátu izolovaných plic |
[119]
|
| 4. | NO není detekovatelný ve vzduchu exhalovaném z izolovaných perfundovaných plic |
[124]
|
| 5. | NO ani jeho oxidační produkt NO2- není detekovatelný v superfuzátu kultury plicních arteriálních endoteliálních buněk |
[125]
|
| 6. | Vzduch z průdušek člověka obsahuje jen minimální množství NO |
[129]
|
Náš nález nejvýše minimální plicní vazokonstrikční odpovědi na akutní podání L-NA a L-NAME [92, 11, 119] přispěl ke značně kontroverzní diskusi. U laboratorních potkanů měla obdobné výsledky naprostá většina laboratoří [89, 90, 110, 91, 111, 113, 138, 114, 115, 116, 117, 139, 140, 141, 142, 120]. Byly však publikovány i opačné výsledky (nezanedbatelná vazokonstrikce) [143, 144]. Podobně většina prací na psech nenalezla plicní vazokonstrikční odpověď na inhibici NO syntázy [112, 143, 145, 146, 147], ale existuje výjimka [148]. U králíka existují údaje pro [149, 150, 151] i proti [152, 153] významné plicní vazokonstrikční odpovědi na deriváty L-argininu. Zřetelná vazokonstrikční odpověď na akutní inhibici NO syntázy byla popsána u ovcí [154, 145, 155], prasat [156, 157, 145] a koček [158]. Ve studiích na lidech je při akutní inhibici NO syntázy zvýšen systémový krevní tlak, tlak v plícnici je nezměněn a srdeční výdej klesá. Následkem toho roste nejen systémový, ale i (podstatně méně) plicní cévní odpor [159, 160]. Akutní vazokonstrikce, vyvolaná v perfundovaných lidských plicích poměrně vysokou dávkou L-NMMA, je jen zčásti inhibovatelná L-argininem [145]. Tyto výsledky proto nedávají dostatečnou představu o tom, do jaké miry je akutní vliv inhibitorů NO syntázy na plicní hemodynamiku lidí důsledkem nespecifických účinků těchto látek.
Zatímco akutní účinek inhibice NO syntázy na plicní cévy je studován velmi často, naše publikace o důsledcích chronické inhibice na plicní oběh [92] dosud zůstavá ojedinělá (přestože chronická inhibice NO syntázy je využívána pro studium úlohy NO v řadě jiných systémů [161, 162, 163, 164]). Obdobné výsledky jiných laboratoří byly publikovány pouze v předběžné formě [165, 166]. Je však pozoruhodné, že v podstatě stejný "pokus" se stejnými výsledky provádí na lidech sama příroda [64]. V těle se souvisle tvoří malé množství derivátů L-argininu, např. L-NMMA a NG,NG-dimetylarginin, které inhibují NO syntázu [167]. Tyto látky jsou normálně rychle vylučovány močí, avšak progresivně se akumulují v plazmě pacientů s chronickým ledvinným selháním [167]. V tomto smyslu jsou tedy tito pacienti v obdobné situaci jako naši pokusní potkani, kterým jsme hladinu inhibitorů NO syntázy v plazmě zvyšovali jejich exogenním podáváním [92]. Tak jako tato naše pokusná zvířata, i tito pacienti mají systémovou hypertenzi a levostrannou srdeční hypertrofii, zatímco plicní hypertenze nepatří mezi komplikace chronického renálního selhání [168, 169]. I když je inhibice NO syntázy jistě nejvýše jedním z mechanismů vzniku systémové hypertenze při chronickém ledvinovém selhání, potvrzují tyto údaje nepřímo validitu našeho pokusu s chronickou inhibicí NO syntázy [92].
Výše diskutované pokusy s akutní inhibicí NO syntázy naznačují, že kromě variability v experimentálních detailech existuje i mezidruhová variabilita, která může odrážet buď rozdílnou úlohu NO v regulaci bazální tenze, anebo rozdílnou citlivost na nespecifické vazokonstrikční působení inhibitorů NO syntázy. Proto je velmi důležité interpretovat tyto nálezy v kontextu ostatních metod. To byl hlavní důvod našich přímých měření produkce NO v plicní cirkulaci [119], respektive v plicním arteriálním endotelu [125], která potvrdila, že je u obou studovaných druhů (potkan a skot) minimální.
Naše výsledky ukazující minimální úlohu NO v regulaci bazálního tonu plicních cév byly potvrzeny studiemi exprese a lokalizace NO syntázy v plicní tkáni. Histochemická a imunohistochemická lokalizace NO syntázy ukázala, že enzym je bohatě přítomen v epitelu dýchacích cest a v endotelu velkých plicních cév potkana, ale chybí v endotelu distálních (odporových) plicních cév [130, 170] (je známo, že pro regulaci plicní hemodynamiky jsou rozhodující menší, rezistentní arterioly). Obsah mRNA pro endoteliální NO syntázu (studovaný in situ hybridizací a reverzní polymerázovou reakcí) je v plicích dospělých potkanu minimální (často pod detekčním limitem) [170, 132]. Dokonce i u dospělých ovcí, (kde byla popsána vazokonstrikční odpověď na inhibitory NO syntázy [154, 145, 155]), je NO syntáza imunohistochemicky detekovatelná pouze v endotelu velkých, avšak nikoliv distálních plicních cév [171]. Údaje o lokalizaci NO syntázy v lidských plicích nejsou jednotné. Giaid a Salehová [172] našli v poměrně rozsahlé studii NO syntázu v endotelu plicních cév zdravých osob. Proti této studii publikovali Xue a Johns [173] metodologické výhrady a sami našli v plicních cévách zdravých lidí jen malé množství NO syntázy; jejich soubor byl však malý.
Je třeba upozornit, že tato diskuse se týká pouze dospělých jedinců. Řada prací ukazuje signifikantní plicní vazokonstrikční odpověď na inhibitory NO syntázy u fétu a novorozence [174, 175, 176, 177, 178, 179, 180, 181, 182, 183] 8, která po narození pozvolna klesá [184, 185] 9. Exprese endoteliální NO syntázy v plicích má obdobný trend [186].
| Předchozí kapitola |
|
8. Reid LM: Structural remodeling of the pulmonary vasculature by environmental change and disease. In: Wagner WW, Weir EK, Eds. The Pulmonary Circulation and Gas Exchange. Armonk, Futura, 1994; 77-110.
11. Hampl V, Weir EK, Archer SL: Endothelium-derived nitric oxide is less important for basal tone regulation in the pulmonary than the renal vessels of adult rat. J Vasc Med Biol 1994; 5: 22-30.
51. Nathan C, Xie Q-W: Regulation of biosynthesis of nitric oxide. J Biol Chem 1994; 269: 13725-13728.
64. Hampl V, Cornfield DN, Huang J, Shultz PJ, Weir EK, Archer SL: Nitric oxide. In: Peacock AJ, Ed. Pulmonary Circulation: A Handbook for Clinicians. London, Chapman and Hall, 1996; 99-114.
65. Hampl V, Archer SL: The role of endogenous nitric oxide in acute hypoxic pulmonary vasoconstriction. In: Zapol WM, Bloch KD, Eds. Nitric Oxide and the Lung. New York, Marcel Dekker, 1997; 113-135.
75. Archer SL, Hampl V: NG-monomethyl-L-arginine causes nitric oxide synthesis in isolated arterial rings: trouble in paradise. Biochem Biophys Res Commun 1992; 188: 590-596. (->abstrakt)
85. Brashers VL, Peach MJ, Rose CE: Augmentation of hypoxic pulmonary vasoconstriction in the isolated perfused rat lung by in vitro antagonists of endothelium-dependent relaxation. J Clin Invest 1988; 82: 1495-1502.
86. Weir EK, Will JA: Oxidants: a new group of pulmonary vasodilators. Bull Eur Physiopathol Respir 1982; 18: 81-85.
87. Garcia JGN, Noonan TC, Jubiz W, Malik AB: Leukotrienes and the pulmonary microcirculation. Am Rev Respir Dis 1987; 136: 131-169.
88. Mazmanian GM, Audet B, Brink C, Cerrina J, Kirkiacharian S, Weiss M: Methylene blue potentiates vascular reactivity in isolated rat lungs. J Appl Physiol 1989; 66: 1040-1045.
89. Archer SL, Tolins JP, Raij L, Weir EK: Hypoxic pulmonary vasoconstriction is enhanced by inhibition of the synthesis of an endothelium derived relaxing factor. Biochem Biophys Res Commun 1989; 164: 1198-1205.
90. Robertson BE, Warren JB, Nye PCG: Inhibition of nitric oxide synthesis potentiates hypoxic vasoconstriction in isolated rat lungs. Exp Physiol 1990; 75: 255-257.
91. Liu S, Crawley DE, Barnes PJ, Evans TW: Endothelium-derived relaxing factor inhibits hypoxic pulmonary vasoconstriction in rats. Am Rev Respir Dis 1991; 143: 32-37.
92. Hampl V, Archer SL, Nelson DP, Weir EK: Chronic EDRF inhibition and hypoxia: effects on pulmonary circulation and systemic blood pressure. J Appl Physiol 1993; 75: 1748-1757. (->abstrakt)
93. Hislop A, Reid L: New findings in pulmonary arteries of rats with hypoxia-induced pulmonary hypertension. Br J Exp Pathol 1976; 57: 542-554.
94. Hampl V, Weir EK, Archer SL: EDRF modulates basal and flow-induced tone in isolated kidneys, but not lungs, of adult rats. Am. Rev. Respir. Dis. 1993; 147: A225.
95. Hauge A: Conditions governing the pressor response to ventilation hypoxia in isolated perfused rat lungs. Acta Physiol Scand 1968; 72: 33-44.
96. McMurtry IF, Davidson AB, Reeves JT, Grover RF: Inhibition of hypoxic pulmonary vasoconstriction by calcium antagonists in isolated rat lungs. Circ Res 1976; 38: 99-104.
97. Hampl V, Herget J: Perinatal hypoxia increases hypoxic pulmonary vasoconstriction in adult rats recovering from chronic exposure to hypoxia. Am Rev Respir Dis 1990; 142: 612-624.
98. Nishiitsutsuji-Uwo JM, Ross BD, Krebs HA: Metabolic activities of the isolated perfused rat kidney. Biochem J 1967; 103: 852-862.
99. Radermacher J, Klanke B, Schurek H-J, Stolte HF, Frolich JC: Importance of NO/EDRF for glomerular and tubular function: studies in the isolated perfused rat kidney. Kidney Int 1992; 41: 1549-1559.
100. Hecker M, Mulsch A, Bassenge E, Busse R: Vasoconstriction and increased flow: two principal mechanisms of shear stress-dependent endothelial autacoid release. Am J Physiol 1993; 265: H828-H833.
101. Bevan JA, Henrion D: Pharmacological implications of the flow-dependence of vascular smooth muscle tone. Annu Rev Pharmacol Toxicol 1994; 34: 173-190.
102. Kuchan MJ, Jo H, Frangos JA: Role of G proteins in shear stress-mediated nitric oxide production by endothelial cells. Am J Physiol 1994; 267: C753-C758.
103. Baylis C, Harton P, Engels K: Endothelial derived relaxing factor controls renal hemodynamics in the normal kidney. J Am Soc Nephrol 1990; 1: 875-881.
104. Radermacher J, Forstermann U, Frolich JC: Endothelium-derived relaxing factor influences renal vascular resistance. Am J Physiol 1990; 259: F9-F17.
105. Tolins JP, Palmer RMJ, Moncada S, Raij L: Role of endothelium-derived relaxing factor in regulation of renal hemodynamic responses. Am J Physiol 1990; 258: H655-H662.
106. Lahera V, Salom MG, Fiksen-Olsen MJ, Romero JC: Mediatory role of endothelium-derived nitric oxide in renal vasodilatory and excretory effects of bradykinin. Am J Hypertens 1991; 4: 260-262.
107. Tolins JP, Raij L: Effects of amino acid infusion on renal hemodynamics. Role of endothelium-derived relaxing factor. Hypertension 1991; 17: 1045-1051.
108. Walder CE, Thiemermann C, Vane JR: The involvement of endothelium-derived relaxing factor in the regulation of renal cortical blood flow in the rat. Br J Pharmacol 1991; 102: 967-973.
109. Gardes J, Poux J-M, Gonzalez M-F, Alhenc-Gelas F, Menard J: Decreased renin release and constant kallikrein secretion after injection of L-NAME in isolated perfused rat kidney. Life Sci 1992; 50: 987-993.
110. Hasunuma K, Yamaguchi T, Rodman DM, O'Brien RF, McMurtry IF: Effects of inhibitors of EDRF and EDHF on vasoreactivity of perfused rat lungs. Am J Physiol 1991; 260: L97-L104.
111. Bhattacharya S, Bhattacharya J: Segmental vascular responses to voltage-gated calcium channel potentiation in rat lung. J Appl Physiol 1992; 73: 657-663.
112. Nishiwaki K, Nyhan DP, Rock P, Desai PM, Peterson WP, Pribble CG, Murray PA: Nomega-nitro-L-arginine and pulmonary vascular pressure-flow relationship in conscious dogs. Am J Physiol 1992; 262: H1331-H1337.
113. Barer G, Emery C, Stewart A, Bee D, Howard P: Endothelial control of the pulmonary circulation in normal and chronically hypoxic rats. J Physiol 1993; 463: 1-16.
114. Oka M, Hasunuma K, Webb SA, Stelzner TJ, Rodman DM, McMurtry IF: EDRF suppresses an unidentified vasoconstrictor mechanism in hypertensive rat lungs. Am J Physiol 1993; 264: L587-L597.
115. Russ RD, Walker BR: Maintained endothelium-dependent pulmonary vasodilation following chronic hypoxia in the rat. J Appl Physiol 1993; 74: 339-344.
116. Russell P, Wright C, Kapeller K, Barer G, Howard P: Attenuation of chronic hypoxic pulmonary hypertension in rats by cyclooxygenase products and by nitric oxide. Eur Respir J 1993; 6: 1501-1506.
117. Zhao L, Crawley DE, Hughes JMB, Evans TW, Winter RJD: Endothelium-derived relaxing factor activity in rat lung during hypoxic pulmonary vascular remodeling. J Appl Physiol 1993; 74: 1061-1065.
118. Grimminger F, Spiestersbach R, Weismann N, Seeger W: Nitric oxide generation and hypoxic vasoconstriction in buffer-perfused rabbit lungs. Am. J. Respir. Crit. Care Med. 1994; 149: A21.
119. Isaacson TC, Hampl V, Weir EK, Nelson DP, Archer SL: Increased endothelium-derived nitric oxide in hypertensive pulmonary circulation of chronically hypoxic rats. J Appl Physiol 1994; 76: 933-940. (->abstrakt)
120. Wanstall JC, Hughes IE, O'Donnell SR: Evidence that nitric oxide from the endothelium attenuates inherent tone in isolated pulmonary arteries from rats with hypoxic pulmonary hypertension. Br J Pharmacol 1995; 114: 109-114.
121. Vallet B, Curtis SE, Winn MJ, King CE, Chapler CK, Cain SM: Hypoxic vasodilation does not require nitric oxide (EDRF/NO) synthesis. J Appl Physiol 1994; 76: 1256-1261.
122. Park KH, Rubin LE, Gross SS, Levi R: Nitric oxide is a mediator of hypoxic coronary vasodilation. Relation to adenosine and cyclooxygenase-derived metabolites. Circ Res 1992; 71: 992-1001.
123. Skinner MR, Marshall JM: Studies on the roles of ATP, adenosine and nitric oxide in mediating muscle vasodilatation induced in the rat by acute systemic hypoxia. J Physiol 1996; 495: 553-560.
124. Hampl V, Tristani-Firouzi M, Hutsell TC, Archer SL: Nebulized nitric oxide/nucleophile adduct reduces chronic pulmonary hypertension. Cardiovasc Res 1996; 31: 55-62. (->abstrakt)
125. Hampl V, Cornfield DN, Cowan NJ, Archer SL: Hypoxia potentiates nitric oxide synthesis and transiently increases cytosolic calcium levels in pulmonary artery endothelial cells. Eur Respir J 1995; 8: 515-522. (->abstrakt)
126. Hampl V, Cowan NJ, Archer SL: Acute hypoxia increases nitric oxide synthesis in cultured pulmonary arterial cells. Am. J. Respir. Crit. Care Med. 1995; 151: A627.
127. Gustafsson LE, Leone AM, Persson MG, Wiklund NP, Moncada S: Endogenous nitric oxide is present in the exhaled air of rabbits, guinea pigs and humans. Biochem Biophys Res Commun 1991; 181: 852-857.
128. Borland C, Cox Y, Higenbottam T: Measurement of exhaled nitric oxide in man. Thorax 1993; 48: 1160-1162.
129. Dillon WC, Hampl V, Shultz PJ, Rubins JB, Archer SL: Origins of breath nitric oxide in humans. Chest 1996; 110: 930-938. (->abstrakt)
130. Xue C, Rengasamy A, Lecras TD, Koberna PA, Dailey GC, Johns RA: Distribution of NOS in normoxic vs. hypoxic rat lung: upregulation of NOS by chronic hypoxia. Am J Physiol 1994; 11: L667-L678.
131. Lundberg JON, Farkas-Szallasi T, Weitzberg E, Rinder J, Lindholm J, Anggard A, Hokfelt T, Lundberg JM, Alving K: High nitric oxide production in human paranasal sinuses. Nature Medicine 1995; 1: 370-373.
132. Shaul PW, North AJ, Brannon TS, Ujiie K, Wells LB, Nisen PA, Lowenstein CJ, Snyder SH, Star RA: Prolonged in vivo hypoxia enhances nitric oxide synthase type I and type III gene expression in adult rat lung. Am J Respir Cell Mol Biol 1995; 13: 167-174.
133. Gerlach H, Rossiant R, Pappert D, Knorr M, Falke KJ: Autoinhalation of nitric oxide after endogenous synthesis in nasopharynx. Lancet 1994; 343: 518-519.
134. Lundberg JO, Weitzberg E, Nordvall SL, Kuylenstierna R, Lundberg JM, Alving K: Primarily nasal origin of exhaled nitric oxide and absence in Kartagener's syndrome. Eur Respir J 1994; 7: 1501-4.
135. Dillon WC, Archer SL, Nelson DP: Nasal origins of breath nitric oxide. Am. J. Respir. Crit. Care Med. 1995; 151: A103.
136. Schedin U, Frostell C, Persson MG, Jakobsson J, Andersson G, Gustafsson LE: Contribution from upper and lower airways to exhaled endogenous nitric oxide in humans. Acta Anaesthesiol Scand 1995; 39: 327-332.
137. Kimberley B, Nejadnik B, Giraud GD, Holden WE: Nasal contribution to exhaled nitric oxide at rest and during breatholding in humans. Am J Respir Crit Care Med 1996; 153: 829-836.
138. Baudouin SV, Evans TW: Action of carbon dioxide on hypoxic pulmonary vasoconstriction in the rat lung: evidence against specific endothelium-derived relaxing factor- mediated vasodilation. Crit Care Med 1993; 21: 740-6.
139. Dumas JP, Dumas M, Sgro C, Advenier C, Giudicelli JF: Effects of two K+ channel openers, aprikalim and pinacidil, on hypoxic pulmonary vasoconstriction. Eur J Pharmacol 1994; 263: 17-23.
140. Feng CJ, Cheng DY, Kaye AD, Kadowitz PJ, Nossaman BD: Influence of Nomega-nitro-L-arginine methyl ester, LY83583, glybenclamide and L158809 on pulmonary circulation. Eur J Pharmacol 1994; 263: 133-140.
141. Fox GA, Paterson NAM, McCormack DG: Effect of inhibition of NO synthase on vascular reactivity in a rat model of hyperdynamic sepsis. Am J Physiol 1994; 36: H1377-H1382.
142. Hassassian H, Burnstock G: Interacting roles of nitric oxide and ATP in the pulmonary circulation of the rat. Br J Pharmacol 1995; 114: 846-850.
143. Barnard JW, Wilson PS, Moore TM, Thompson WJ, Taylor AE: Effect of nitric oxide and cyclooxygenase products on vascular resistance in dog and rat lungs. J Appl Physiol 1993; 74: 2940-2948.
144. Roos CM, Frank DU, Xue C, Johns RA, Rich GF: Chronic inhaled nitric oxide: effects on pulmonary vascular endothelial function and pathology in rats. J Appl Physiol 1996; 80: 252-60.
145. Cremona G, Wood AM, Hall LW, Bower EA, Higenbottam T: Effect of inhibitors of nitric oxide release and action on vascular tone in isolated lungs of pig, sheep, dog and man. J Physiol 1994; 481: 185-195.
146. Hakim TS: Flow-induced release of EDRF in the pulmonary vasculature: site of release and action. Am J Physiol 1994; 267: H363-H369.
147. Leeman M, De Beyl VZ, Delcroix M, Naeije R: Effects of endogenous nitric oxide on pulmonary vascular tone in intact dogs. Am J Physiol 1994; 266: H2343-H2347.
148. Perrella MA, Hildebrand FL, Margulies KB, Burnett JC: Endothelium-derived relaxing factor in regulation of basal cardiopulmonary and renal function. Am J Physiol 1991; 261: R323-R328.
149. Persson MG, Gustafsson LE, Wiklund NP, Moncada S, Hedqvist P: Endogenous nitric oxide as a probable modulator of pulmonary circulation and hypoxic pressor response in vivo. Acta Physiol Scand 1990; 140: 449-457.
150. Wiklund NP, Persson MG, Gustafsson LE, Moncada S, Hedquist P: Modulatory role of endogenous nitric oxide in pulmonary circulation in vivo. Eur J Pharmacol 1990; 185: 123-124.
151. Persson MG, Kalzen H, Gustafsson LE: Oxygen or low concentrations of nitric oxide reverse pulmonary vasoconstriction induced by nitric oxide synthesis inhibition in rabbits. Acta Physiol Scand 1994; 150: 405-411.
152. Xu H, Pearl RG: Effects of L-glutamine on pulmonary hypertension in the perfused rabbit lung. Pharmacology 1994; 48: 260-264.
153. Grimminger F, Spriestersbach R, Weissmann N, Walmrath D, Seeger W: Nitric oxide generation and hypoxic vasoconstriction in buffer-perfused rabbit lungs. J Appl Physiol 1995; 78: 1509-1515.
154. Meyer J, Lentz CW, Herndon DN, Nelson S, Traber LD, Traber DL: Effects of halothane anesthesia on vasoconstrictor response to NG-nitro-L-arginine methyl ester, an inhibitor of nitric oxide synthesis, in sheep. Anesth Analg 1993; 77: 1215-1221.
155. Koizumi T, Gupta R, Banerjee M, Newman JH: Changes in pulmonary vascular tone during exercise: effects of nitric oxide (NO) synthase inhibition, L-arginine infusion, and NO inhalation. J Clin Invest 1994; 94: 2275-2282.
156. Alving K, Fornhem C, Lundberg JM: Pulmonary effects of endogenous and exogenous nitric oxide in the pig: relation to cigarette smoke inhalation. Br J Pharmacol 1993; 110: 739-746.
157. van Gelderen EM, Den Boer MO, Saxena PR: NG-nitro l-arginine methyl ester: systemic and pulmonary haemodynamics, tissue blood flow and arteriovenous shunting in the pig. Naunyn-Schmiedeberg's Arch Pharmacol 1993; 348: 417-423.
158. McMahon TJ, Hood JS, Bellan JA, Kadowitz PJ: Nomega-nitro-L-arginine methyl ester selectively inhibits pulmonary vasodilator responses to acetylcholine and bradykinin. J Appl Physiol 1991; 71: 2026-2031.
159. Stamler JS, Loh E, Roddy M-A, Currie KE, Creager MA: Nitric oxide regulates basal systemic and pulmonary vascular resistance in healthy humans. Circulation 1994; 89: 2035-2040.
160. Blitzer ML, Loh E, Roddy MA, Stamler JS, Creager MA: Endothelium-derived nitric oxide regulates systemic and pulmonary vascular resistance during acute hypoxia in humans. J Am Coll Cardiol 1996; 28: 591-596.
161. Arnal J-F, Warin L, Michel J-B: Determinants of aortic cyclic guanosine monophosphate in hypertension induced by chronic inhibition of nitric oxide synthase. J Clin Invest 1992; 90: 647-652.
162. Bank N, Aynedjian HS, Khan G: Mechanism of vasoconstriction induced by chronic inhibition of nitric oxide in rats. Hypertension 1994; 24: 322-328.
163. Navarro J, Sanchez A, SŠiz J, Ruilope LM, Garcia-Estan J, Romero JC, Moncada S, Lahera V: Hormonal, renal, and metabolic alterations during hypertension induced by chronic inhibition of NO in rats. Am J Physiol 1994; 36: R1516-R1521.
164. Pechanova O, Bernatova I: Effect of long-term NO synthase inhibition on cyclic nucleotide content in rat tissues. Physiol Res 1996; 45: 305-309.
165. Emery C: Effect of chronic oral treatment with L-nitro arginine methyl ester on hypoxic vasoconstriction in rat lung. Am Rev Respir Dis 1993; 147 (suppl.): A226.
166. Kouyoumdjian C, Adnot S, Carville C, Eddahibi S, Raffestin B: Effects of short term and long term inhibition of nitric oxide (NO) synthesis in normoxic and hypoxic rats. Am Rev Respir Dis 1993; 147 (suppl.): A494.
167. Vallance P, Leone A, Calver A, Collier J, Moncada S: Accumulation of an endogenous inhibitor of nitric oxide synthesis in chronic renal failure. Lancet 1992; 339: 572-575.
168. Bartges JW, Willis AM, Polzin DJ: Hypertension and renal disease. Vet Clin North Am Small Anim Pract 1996; 26: 1331-45.
169. Kovac A, Demes M, Pavlovic M, Slugen I: Chronická renalna insuficienia: soucasny pohl'ad na patogenezu, klinicky obraz a liecbu. Vnitr Lek 1996; 42: 49-53.
170. Kawai N, Bloch DB, Filippov G, Rabkina D, Suen H-C, Losty PD, Janssens SP, Zapol WM, De la Monte S: Constitutive endothelial nitric oxide synthase gene expression is regulated during lung development. Am J Physiol 1995; 12: L589-L595.
171. Halbower AC, Tuder RM, Franklin WA, Pollock JS, Forstermann U, Abman SH: Maturation-related changes in endothelial nitric oxide synthase immunolocalization in developing ovine lung. Am J Physiol 1994; 11: L585-L591.
172. Giaid A, Saleh D: Reduced expression of endothelial nitric oxide synthase in the lungs of patients with pulmonary hypertension. N Engl J Med 1995; 333: 214-221.
173. Xue C, Johns RA: Endothelial nitric oxide synthase in the lungs of patients with pulmonary hypertension [letter]. N Engl J Med 1995; 333: 1642-1644.
174. Abman SH, Chatfield BA, Hall SL, McMurtry IF: Role of endothelium-derived relaxing factor during transition of pulmonary circulation at birth. Am J Physiol 1990; 259: H1921-H1927.
175. Davidson D, Eldemerdash A: Endothelium-derived relaxing factor: evidence that it regulates pulmonary vascular resistance in the isolated neonatal guinea pig lung. Pediatr Res 1991; 29: 538-542.
176. Fineman JR, Heymann MA, Soifer SJ: Nomega-nitro-L-arginine attenuates endothelium-dependent pulmonary vasodilation in lambs. Am J Physiol 1991; 260: H1299-H1306.
177. Chang J-K, Moore P, Fineman JR, Soifer SJ, Heymann MA: K+ channel pulmonary vasodilation in fetal lambs: role of endothelium-derived nitric oxide. J Appl Physiol 1992; 73: 188-194.
178. Moore P, Velvis H, Fineman JR, Soifer SJ, Heymann MA: EDRF inhibition attenuates the increase in pulmonary blood flow due to oxygen ventilation in fetal lambs. J Appl Physiol 1992; 73: 2151-2157.
179. Shaul PW, Farrar MA, Zellers TM: Oxygen modulates endothelium-derived relaxing factor production in fetal pulmonary arteries. Am J Physiol 1992; 262: H355-H364.
180. Fineman JR, Wong J, Morin FC, Wild LM, Soifer SJ: Chronic nitric oxide inhibition in utero produces persistent pulmonary hypertension in newborn lambs. J Clin Invest 1994; 93: 2675-2683.
181. Kinsella JP, Ivy DD, Abman SH: Ontogeny of NO activity and response to inhaled NO in the developing ovine pulmonary circulation. Am J Physiol 1994; 36: H1955-H1961.
182. Wang Y, Coceani F: EDRF in pulmonary resistance vessels from fetal lamb: stimulation by oxygen and bradykinin. Am J Physiol 1994; 266: H936-H943.
183. Nelin LD, Thomas CJ, Dawson CA: Effect of hypoxia on nitric oxide production in neonatal pig lung. Am J Physiol 1996; 271: H8-14.
184. Perreault T, De Marte J: Maturational changes in endothelium-derived relaxations in newborn piglet pulmonary circulation. Am J Physiol 1993; 264: H302-H309.
185. Shaul PW, Farrar MA, Magness RR: Pulmonary endothelial nitric oxide production is developmentally regulated in the fetus and newborn. Am J Physiol 1993; 265: H1056-H1063.
186. North AJ, Star RA, Brannon TS, Ujiie K, Wells LB, Lowenstein CJ, Snyder SH, Shaul PW: Nitric oxide synthase type I and type III gene expression are developmentally regulated in rat lung. Am J Physiol 1994; 266: L635-L641.
| Předchozí kapitola |
|
6 Redukce NO3- nebylo třeba, protože perfuzát (Krebsův roztok s albuminem) neobsahoval oxyhemoproteiny (kap. 3.1).
7 Rozdělovací koeficient 20 znamená, že 20x více NO bude přítomno v plynné fázi nad hladinou roztoku než bude rozpuštěno v roztoku.
8 Většina těchto prací byla ovšem provedena na ovcích, kde i v dospělosti působí inhibitory NO syntázy plicní vazokonstrikci [154, 145, 155].
9 Shaul a spol. [185] vyvodili ze svých pokusů závěr, že bazální syntéza NO v plicních cévách po narození stoupá. Tento závěr byl ovšem založen na pokusech provedených za nefysiologicky vysokého PO2 (680 mmHg). Jejich data získaná za normoxie (PO2 = 150) svědčí o opaku.
| Předchozí kapitola |
|